Pertumbuhan Rumput Dengan Penambahan Mikro Organisme Tanah

PERTUMBUHAN RUMPUT DENGAN PENAMBAHAN MIKROORGANISME TANAH
Eva Kurniawati, Iwan Kurniawan, Nando suito Sihombing
BALAI PEMBIBITAN TERNAK UNGGUL DAN HIJAUAN PAKAN TERNAK SIBORONGBORONG
ABSTRAK
Ketersediaan pakan ternak, khususnya hijauan masih merupakan masalah yang sulit diatasi terutama pada musim kemarau. Lahan untuk hijauan pakan ternak juga semakin berkurang karena tergeser oleh penggunanan lahan pemukiman dan tanaman pangan. Pada saat sekarang ini sulit untuk mendapatkan lahan yang subur untuk tanaman pakan ternak, karena lahan yang subur pada umumnya digunakan untuk tanaman pangan sehingga tanaman pakan ternak ditanam dengan memanfaatkan lahan marginal. Penggunaan mikroorganisme tanah bernilai positif terhadap pertumbuhan rumput.
Kata kunci : pertumbuhan rumput, mikroorganisme tanah
GRASS GROWTH WITH THE ADDITION OF LAND MICROORGANISM
ABSTRACT
The availability of animal feed, especially forage is still a difficult problem to overcome, especially in the dry season. Land for forage forage is also increasingly reduced because it is displaced by the use of residential land and food plants. At the present time it is difficult to get fertile land for animal feed plants, because fertile land is generally used for food crops so that livestock feed crops are planted using marginal land. The use of soil microorganisms is positive for grass growth.
Keywords: Grass Growth, Soil Microorganisms
Pendahuluan
Pada saat sekarang ini sulit untuk mendapatkan lahan yang subur untuk tanaman pakan ternak, karena lahan yang subur pada umumnya digunakan untuk tanaman pangan sehingga tanaman pakan ternak ditanam dengan memanfaatkan lahan marginal, lahan yang kurang dengan unsur hara.
Salah satu upaya menyiasati lahan marginal yang kurang akan unsur hara ialah dengan menggunakan Mikoriza Arbuskula (FMA). FMA ini berperan untuk melepaskan ikatan P dari mineral liat dan menyediakannya bagi tanaman. Tanaman yang bermikoriza biasanya lebih tahan kering daripada yang tidak bermikoriza. Kekeringan yang menyebabkan rusaknya jaringan korteks, kemudian matinya perakaran pengaruhnya tidak akan permanen pada akar yang bermikoriza. Akar yang bermikoriza akan cepat kembali pulih setelah periode kekurangan air berlalu. Hal ini disebabkan oleh hifa Fungi mampu menyerap air pada pori-pori tanah, pada saat akar tanaman sudah tak mampu. Selain itu penyebaran hifa didalam tanah sangat luas, sehingga dapat mengambil air relatif banyak.tanaman bermikoriza juga tahan tehadap serangan patogen akar. Mikoriza menggunakan hampir semua mikroba tanah lain yang berperan didalam penyedian unsur hara adalah mikroba pelarut fosfat (P)dan Kalium (K).
Kajian Pustaka
Kriteria Tanah Ultisol
Ultisol umumnya tanah ber pH rendah yang bereaksi masam, produktifitasnya rendah, kapasitas tukar kation (KTK) dan kejenuhan basa (KB) yang randah kejenuhan alumunium (AL) yang tinggi, kandungan bahan organik rendah dan peka terhadap erosi. Masalah utama pada tanah ultisol ini adalah jumlah kelarutan dan kejenuhan AL yang tinggi sehingga mengakibatkan fosfor (P)membentuk senyawa yang tidak larut dengan Al. Akibatnya ketersedian P sangat rendah bagi tanaman sehinggapertumbuhan tanaman terganggu (Sanchez,1992).
Kekahatan P ditanah ultisol merupakan masalah keharaan yang paling penting, sebab kekahatan P tidaklah semata-mata karena kandungan P tanah yang memang rendah akan tetapi juga karena sebagina besar P dalam keadaan tersemat (Hardjowigeno, 1993).
Lynch (1983) menyatakan bahwa teknologi tanah yang dikombinasikan dengan praktek-praktek usaha tani merupakan alat yang sangat penting untuk mengembangkan pertanian pda tanah mineral masam tropika. Teknologi ini mencakup segala upaya memanipulasi jasad renik tanah dan proses metabolik mereka untuk mengoptimumkan produksi tanaman. Penggunaan jasad renik mikoriza telah mulai diupayakan dalam kebijaksanaan pengelolaan tanah mineral masam tropika. Widada dan Kabirun (1995) menemukan bahwa mikoriza mempunyai peranan yang besar dalam pengelolaan tanah mineral masam tropika.
Pada tanah-tanah tersebut ditemukan beberapa spesies mikoriza yang mempunyai ketahanan tinggi terhadap kemasaman dan keracunan AL serta berpotensi besar dalam meningkatkan pertumbuhan dan produksi tanaman.
Mikroorganisme Tanah (Fungi Mikororiza Arbuskula) dan Pengaruhnya terhadap Ketersediaan Serapan Hara dan Pertumbuhan Tanaman.
Asosiasi simbiotik antara jamur dan system perakaran tanaman tinggi diistilahkan dengan mikoriza. Dalam hal ini jamur menginfeksi dan mengkoloni akar tanpa menimbulkan nekrosis sebagaimana biasa terjadi pada infeksi jamur pathogen dan mendapat pasokan nutrisi secara teratur dari tanaman (Rao, 1994).
Istilah mikoriza yang berarti jamur akar pertama kali dikenalkan oleh Frank pada tahun 1855. Hubungan simbiosis antara sistem perakaran tanaman dengan kelompok Fungi tertentu saling menguntungkan. Tanaman mendapat hara lebih banyak sedangkan bagi Funginya sendiri diuntungkan karena mendapat fotosintat dari inangnya. Tanaman bersimbiosis dengan CMA dapat memperbaiki pertumbuhan tanaman dengan meningkatkan penyrapan unsur hara P (Fitter and Hay, 1991). Tanah- tanah yang mempunyai kandungan unsur P minimum merupakan pembatas bagi pertumbuhan tanaman, tetapi dapat menginokulasi CMA tersebut ke dalam tanah akan sangat membantu dalam penyerapan unsur -unsur hara yang diperlukan.
Bagian yang penting dari mikoriza adalah miselium yang terdapat diluar akar, dimana bagian luar hifa tersebut berperan dalam penyerapan unsur hara tanaman. Jarak yang ditempuh oleh hara tanaman dengan adanya mikoriza dapat diperpendek.
Anatomi sederhana dari CMA dapat di lihat pada gambar 1.
Gambar 1. Penampang longitudinal akar yang terinfeksi CMA
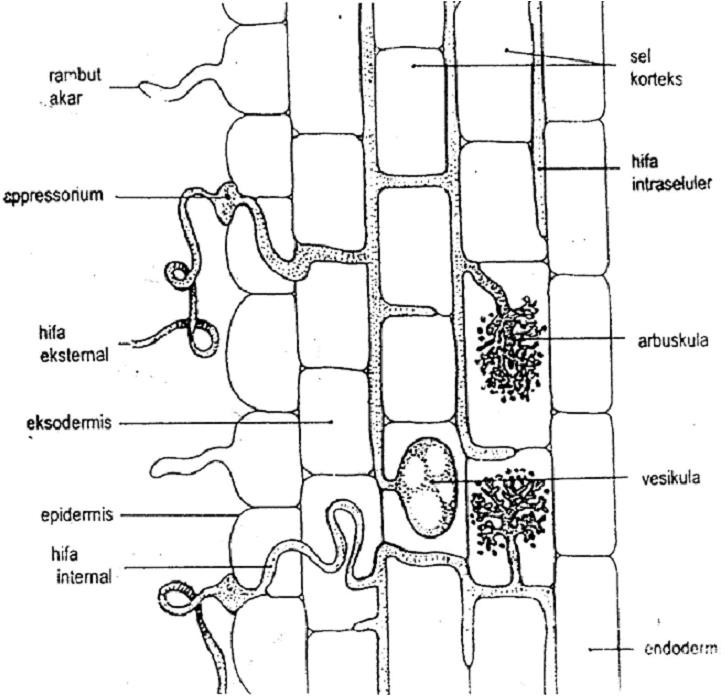
(Sumber : Brundrett et al., 1994)
Hetrick (1984) menyimpulkan bahwa kolonisasi akar dan produksi spora dipengaruhi oleh dua faktor yaitu : spesies Fungi dan lingkungan. Faktor spesies Fungi dibedakan menjadi faktor kerapatan inokulum dan persaingan antar spesies Fungi. Peningkatan kadar nokulum dapat meningkatkan persentase kolonisasi akar sampai titik optimum tertentu (Daft dan Nicolson, 1979).
Keanekaragaman CMA tidak mengikuti pola keanekaragaman tanaman, dan tipe CMA mungkin mengatur keaneka ragaman spesies tanaman (Allen et al., 1995). Adanya simbiosis mutualistik antara CMA dengan perakaran tanaman dapat membantu petumbuhan tanaman menjadi lebih baik, terutama pada tanah-tanah marjinal. Hal ini disebabkan CMA efektif dalam meningkatkan penyerapan unsur hara dan mikro (Allsopp dan Stock, 1993) meningkatkan daya tahan tanaman terhadap serangan pathogen (Wani et al., 1991), meningkatkan ketahan terhadap kekeringan (Devies et al., 1992) dan dapat membantu pertumbuhan tanaman pada daerah yang tercemar logam berat (Munyanziza et al., 1997).
Spesies dan strain CMA mempunyai perbedaan dalam kemampuannya meningkatkan penyerapan hara dan pertumbuhan tanaman (Daniels dan Menge, 1981). Menurut Abbot dan Robson (1992) setiap spesies CMA mempunyai innate effectiveness atau kemempanan spesifik. Keefektivan (effectiveness) diartikan sebagai kemampuan CMA dalam meningkatkan pertumbuhan tanaman pada kondisi tanah yang kurang menguntungkan.
Jenis Tanaman
Jenis tanaman yang berbeda umumnya akan menunjukkan reaksi yang berlainan terhadap infeksi mikoriza dan secara tak langsung mempengaruhi perkembangan infeksi dan kolonisasi jamur mikoriza.
Perbedaan reaksi tanaman itu sangat dipengaruhi oleh aras kepekaan tanaman terhadap infeksi dan sifat ketergantungan tanaman pada mikoriza dalam serapan hara, khususya di tanah kahat P yang mana kedua sifat itu ada kaitannya dengan tipe perakaran dan keadaan fisiologi tanaman. Struktur akar disebutkan Mosse (1981), mempengaruhi tanggapan tanaman terhadap infeksi. Selanjutnya Moose (1981) mencontohkan rerumputan yang panjang total akarnya kali lipat dibandingkan akar tanaman legum diketahui bersifat tergantung pada mikoriza untuk pertumbuhannya. Kebutuhan tanaman akan unsur P, secara fisiologis mempengaruhi kepekaan terhadap infeksi. Tanaman yang kebutuhan P-nya tinggi biasanya lebih peka dan tanggap.
Kualitas Hijauan
Protein kasar merupakan salah satu unsur tanamanyang lebih umum diukur sehubungan dengan kepentingan untuk konsumsi ternak. Rata-rata kandungan protein kasar tanaman menunjukkan bahwa semakintinggi pemberian nitrogen, semakin baik kandungan protein kasar tanaman. Naiknya kandungan protein kasar tersebut mudah diterangkan karena nitrogen merupakan penyusun protein (Mappaona, 1986). Umumnya kandungan protein kasar leguminosa 25% lebih tinggi daripada rumput dengan umur dan pertumbuhan yang sama dan hanya menunjukkan sedikit fluktuasi dari proses pertumbuhan (aminah et al.,1989. Kandungan protein dari rumput berkisar antara 8-17% dan legum antara 15-28% (Wan Mohamed et al., 1987).
Prawiranata et al. (1981) menyatakan bahwa bertambahnya usia tanaman menghasilkan perbandingan daun dengan batang semakin kecil. Kecilnya perbandingan daun dengan batang berpengaruh terhadap protein kasar. Kandungan energi dan protein pakan sangat berperanan terhadap produksi ternak.
Wan Hassan (1987) melaporkan bahwa rumput-rumput yang ada di daerah tropika (salah satunya rumput asli atau rumput yang telah mengalami perbaikan) umumnya mempunyai metabolisme energi antara 7-11 MJ/kg BK pada umur pemotongan antara umur 2-8 minggu, dan energi konsentrasi dari hijauan alami berkisar 7.1-10.1 MJME/kg BK. Banerjee (1978) menyatakan bahwa pakan yang mengandung serat kasar tinggi pada umumnya rendah kandungan energi dan proteinnya. Wilsn (1982) menyatakan bahwa naungan dapat mempengaruhi nilai nutrisi pada pastura, antara lain: rendahnya tingkat karbohidrat terlarut tanaman, meningkatnya kandungan silika dan lignifikasi, menurunnya kecernaan dinding sel, menurunya proporsi jaringan mesophil yang mudah dicrna terhadap jaringan epidermis yang sulit dicerna, meningkatnya persentase kandungan air dari jaringan yang sapat menurunkan konsumsi hijauan oleh ternak, dan protein kasar terkadang lebih tinggi pada tanaman yang berada di bawah naungan.
Pembahasan
Pertumbuhan rumput dalam hal produksi bahan segar rumput Panicum maximum dengan penambahan mikroorganisme tanah meningkat. Ha ini sesuai dengan pernyataan Setiadi (1990) yang menyatakan bahwa tanaman yang bermikoriza akan tumbuh lebih baik dari tanaman tanpa mikoriza, karena mikoriza secara efektif dapat meningkatkan penyerapan unsur hara makro. Selain itu akar yang bermikoriza dapat menyerap unsur hara dalam bentuk terikat dan tersedia bagi tanaman.
Dengan penambahan mikro- organisme tanah juga selaras dengan pertambahan tinggi tanaman. Mikoriza dapat meningkatkan pertumbuhan dan perkem- bangan tanaman melalui simbiosis mutualisme antara mikoriza pada akar tanaman dengan tanaman itu sendiri.
Mikoriza memberikan unsur-unsur hara yang dibutuhkan oleh tanaman untuk tumbuh dan berproduksi, sebaliknya Mikoriza mendapatkan makanan melalui akar tanaman itu sendiri. Hal ini sesuai dengan pernyataan Santoso et al., (2006) yang menyatakan bahwa Karena cuma inang yang berfotosintesa, sebagai imbalannya, sebagian hasil fotosintat (berupa karbohidrat cair) yang dimasak pada daun berklorofil didistribusikan ke bagian akar inang, dan tentunya mikoriza dijaringan korteks akar inang mendapatkan aliran energi untuk hidup dan berkembangbiak di dalam tanah. Dari kegiatan barter antara mikoriza dan inang, maka proses simbiosis mutualistis berlangsung terus menerus dan salin menguntungkan seumur hidup inang.
Pengaruh FMA terhadap pertumbuhan secara umum dinyatakan bahwa tanaman yang bermikoriza tumbuh lebih baik dari tanaman tanpa bermikoriza. Penyebab utama adalah mikoriza secara efektif dapat meningkatkan unsur hara, baik unsur hara makro maupun mikro. Selain daripada itu akar yang bermikoriza dapat menyerap unsur hara dalam bentuk terikat dan tidak tersedia bagi tanaman. Mose (1981) menyebutkan indikasi tanggap tanaman pada serapan P terhadap FMA adalah kandungan P tanaman bermikoriza umumnya lebih tinggi daripada tanaman tanpa mikoriza dan ada kesurupan pengaruh inokulasi FMA dengan pemberian pupuk P terhadap pertumbuhan tanaman di tanah dengan P tersedia rendah sampai sedang. Salain daripada membentuk hifa interna, mikoriza juga membentuk eksterna. Pada hifa eksterna akan terbentuk spora yang merupakan bagaian penting dari mikoriza yang berda di luar kar. Fungsi utama dari hifa ini adalah untuk menyerap fosfor di dalam tanah. P yang terakumulasi pada hifa eksternal dipindahkan ke miselium internal oleh suatu sistem transport dan kemudian ke dalam jaringan tanaman inang.
Semakin banyak dosis mikoriza yang diberikan pada tanaman, maka akan semakin besar pula bintil akar yang dihasilkan oleh tanaman inangnya tersebut. Ini disebabkan karena fungi mikoriza arbuskula bekerja pada akar inangnya seperti jaringan hifa yang semakin banyak. Hal ini sesuai dengan pernyataan Wicaksono dan Ricky (2010), yang menyatakan bahwa mikoriza memberikan manfaat bagi tanaman diantaranya adalah: 1) meningkatkan serapan unsur hara, 2) meningkatkan ketahanan terhadap kekeringan, 3) kerusakan jaringan korteks akibat kekeringan pada perakaran bermikoriza tidak bersifat permanen, 4) memperluas penyebaran hifa dalam tanah sehingga dapat mengambil air relatif lebih banyak, serta 5) memproduksi hormone dan zat pengatur tumbuh seperti auxin, sitokinin, giberelin dan vitamin bagi inangnya.
Kesimpulan dan Saran
Dari pembahasan diatas bisa disimpulkan bahwa mikroorganisme tanah dalam hal ini digunaakan mikoriza arbuskula untuk pertumbuhan rumput hijauan pakan ternak berpengaruh baik untuk pertumbuhan dan produksi rumput sehingga kita tidak hanya dapat memenuhi kebutuhan pakan ternak (rumput) dalam segi kuantitasnya saja tetapi beserta kualitas dari rumput itu sendiri.
Daftar Pustaka
Allen EB, MF. Allen, DJ. Helm, JM. Trape, R. Molina, dan E. Rincon.1995. pattern and regulation of mycorrhizal plant and fungal diversity. Plant and soil. 170 (1) : 47-62.
Allsopp N dan Stock WD.1993. mycoorhizal and seedling growth of slow growing scleropyllis from nutrient poor environments. Acta aoecologica international Journal of Ecology. 14 (5) :577-587.
Banerjee GC. 1978. A Text Book of Animal Husbandry . Fouth Edition. Bombay : Oxford & Publishing Co. New Delhi.
Brundrett MC, Melville L dan Peterson L. 1994. Pratical methods in mycoorhiza research. Mycologue Publications. Ontario, canada. 161 hal.
Daft MJ dan Nicolson TH. 1979. Effects of salinity on tomato. Scientia Hort. 78:83-125.
Daniels BA dan Menge JA. 1981. Evaluation of commercial potential of the VAM fungus, Glomus epigaeus. New Phytol 87 : 345-353.
Daniels B.A dan trappe JM.1980. factors affecting spore germination of vesicular arbuscular mycorrhizal fungus, Glomus epigaeus. Mycologi 72: 457-463.
Devies FT, Potter JR dan Linderman RG. 1992 Mycorrhiza and repeated drought exposure affect drought resistance and extradical hypae development of pepper plants independent of plant size and nutrient content. Journal of plant Physiology. 139 (3) :289-294.
Fitter, AH dan RKM. Hay.1991. Environmental Physiology of plants. Terjemahan dari Sri Handayani, E.D Purbayati dan B. Sri Gandono. Fisiologi Lingkungan Tanaman. Gajah Mada University Press. Yogyakarta.
Horne PM, Stur WW.1999. Mengembangkan Teknologi Hijaun Makanan Ternak bersama Petani Kecil. ACIAR Proc 65 :70-76.
Mappaona. 1986. Pengaruh Naungan dan Pemberian Nitrogen terhadap Produksi Bahan Kering dan Komposisi Kimia Rumput Brachiaria decumbers. Bogor : Program Pascasarjana IPB.
Mc. Ilroy, R.J. 1976. Pengantar Budiday padang Rumput Tropika. Susetyo, penerjemah. Jakarta: Pradyana, Paramita.
Mosse. B. 1981. Vesicular Arbuscular Mycoorhizal Research for Tropical Agricultural Research. Ress. Bull. Hawai. Ins. Trop. Agric. And Human Resources.82 p.
Munyanziza E, Kehri HK dan Bagyaraj DJ.1997. Agricultural Intensification, Soil biodiorversity and agro-ecosystem function in the tropic : the role of mycorrhiza in crops and tress. Applied Soil Ecology. 6 (1) :77-85.
Prawiranata W, harran S, tjondronegoro P. 1981. Dasar-dasar Fisiologi Tumbuhan, Bogor: Departemen Botani, Fakultas Pertanian, IPB.
Reksohadiprodji S. 1994. Produksi tanaman Hijauan makanan Ternak/Tropika. Yogyakarta : BPFE.
Sanchez, P.A.1992. Sifat dan Pengelolaan Tanah Tropika. Terjemahan Amir Hamzah. ITB Bandung. 397 hal.
Santosa D.A. 1986. Teknik dan Metode Penelitian ikoriza Vesicular Arbuscular. Lab Biologi Tanah. IPB. Bogor.57 hal.
Setiadi Y. 1991. Pemanfaatan Mikroorganisme dalam Kehutanan. Departemen P dan K. Dirjen. DIKTI PAU Bioteknologi IPB Bogor.
Wani S.P, Mc Gill W.B and Tewari J.P. 1991. Mycorrhizal and Common Root infection and Nutrient Accumulation in Barley Grown on Breton Loam using N from Biological Fixation or Fertilizer. Biology and Fertility of Soil. 12(1) : 46-54.
Widada, J dan S. Kabirun. 1995. Peran Mikoriza Arbuskular dalam Pengelolaan Tanah Mineral Masam Tropika. Dalam Prosiding Penatagunaan tanah sebagai Perangkat Penataan Ruang dalam Rangka Meningkatkan Kesejahteraan Rakyat. Konges nasional VI HITI, Serpong.
Wan Hasan. 1987. Utilization of Gound Vegetation for Animal Rearing. Proc of The Rubber Research Institude of Malaysia Planters Conference, 17-19 October 1987, Kuala Lumpur : 265-272.
Wan Mohammed, Hutagalung WE, Chen CP. 1987 Feed Availability, Utilization and Constraints in plantations of Asia and The Pasific : Performence and Prospect. Tropical Grasslands 21 : 159-186.
Wilson JR. 1982. Environmental and nutritional factor affecting herbage quality. United Kingdom. P.111-131.



